

Abstract
Background
Histones are nuclear proteins that can be released extracellularly following cell death or certain mechanisms of cell activation. Infusion of high doses of histones into mice causes lethality. Circulating levels of extracellular histones are elevated in several disease states associated with inflammation and thrombosis. The mechanisms by which extracellular histones induce morbidity and mortality are not fully understood. Histones may activate or be toxic to several cell types in vitro. Most of these activities have been shown to be dependent on histone signaling through toll-like receptors (TLRs). Red blood cells (RBC) do not express TLRs. However, previous studies have reported that histones promote RBC aggregation, increase shear-dependent RBC fragility, reduce deformability under flow and cause hemolytic anemia in mice. Histones also induce calcium influx and promote PS expression on RBC membranes thereby supporting the prothrombinase activity and shortening clotting times in plasma. Histones may insert into cell membranes (independently of TLRs), and form pore-like structures similar to ionophore. The role of this process in histone-induced RBC toxicity has not been fully investigated.
Objective
We assessed the ability of individual histones to promote the release of extracellular hemoglobin (Ex-Hb) and RBC-derived microvesicles (RBC-MVs), as well as RBC-MVs' ability to initiate or propagate thrombin generation (TG) in MV-free plasma.
Methods
RBCs were isolated from citrated whole blood from healthy volunteers, and were re-suspended in buffer with or without 10uM A23187 (calcium ionophore), recombinant histones H2A, H2B, H3 or H4 for 1 hour at 37°C.
To assess histone binding, RBCs were washed and dual-stained with fluorophore-conjugated anti-glycophorin A (GPA) and anti-panhistone antibodies. Stained RBCs were analyzed using imaging flow cytometry, and were considered positive for histone binding if the fluorescence intensity was > 95th percentile of RBCs processed in the absence of histones.
RBCs were removed by centrifugation at 2,500g for 15 minutes x2. RBC-MVs present in the supernatant were pelleted by centrifugation at 20,000g for 30 min. Isolated RBC-MVs were then re-suspended in buffer to assess PS exposure using the microparticle activity assay (Hyphen Biomed). RBC-MVs were also re-suspended in re-calcified MV-free plasma supplemented with 4uM 41%PC/44%PE/15%PS lipid vesicles to assess their ability to initiate TG by monitoring the activity of thrombin on a fluorogenic substrate over time. To assess their ability to propagate TG, coagulation was initiated by 1pM tissue factor in MV-free plasma containing histone-induced RBC-MVs without addition of synthetic lipid vesicles. In this condition, phospholipids present on the surface of histone-induced RBC-MVs are the limiting factor for TG.
The supernatant collected after pelleting RBC-MVs was used to measure Ex-Hb by absorbance at 540 nm.
Results
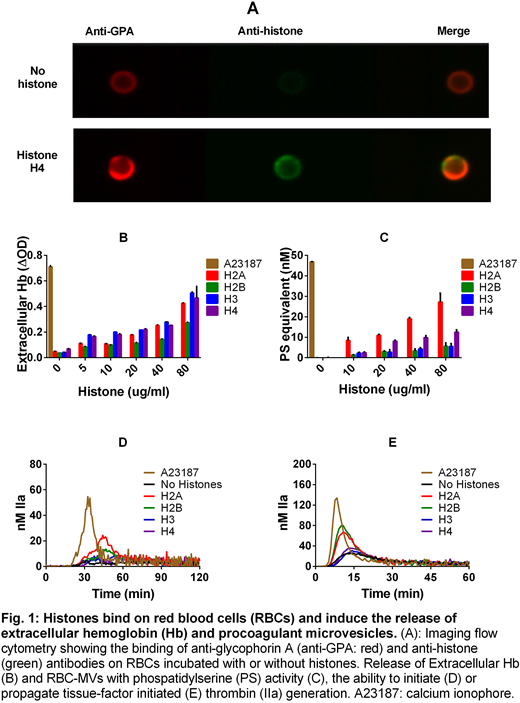
The percentage of RBCs positive for histone binding was 21.6%, 2.3%, 76.9% and 98.1% for H2A, H2B, H3 and H4, respectively. Representative images of RBC staining for GPA and histone H4 is shown in Figure 1A.
All histone types induced the release of Ex-Hb (Fig. 1B) and RBC-MVs that possess PS activity (Fig. 1C) in a dose-dependent manner, with the highest PS activity observed with H2A. RBC-MVs released by H2A or H2B clearly initiated TG in MV-free normal pool plasma (Fig.1D). They also significantly enhanced TF-initiated TG in the absence of additional lipid vesicles. The release of Ex-Hb and procoagulant RBC-MVs induced by histones were re-capitulated by ionophore to a greater degree, as expected (Fig.1B-E).
Conclusions
All 4 types of core histones bind to RBCs and induce the release of Ex-Hb. Our findings suggest that despite their lower binding to RBCs, histones H2A and H2B may have a greater ability to induce the release of procoagulant RBC-MVs that can initiate and propagate TG in plasma. The reason for the differential toxicity of histone types on RBCs and the discrepancy between the relative effect of histone-induced RBC-MVs on PS equivalents and TG are unknown. Histone-induced RBC toxicity appears to mimic calcium ionophore, supporting the idea that histones may insert and form functional pores in cell membranes. The contribution of Ex-Hb and procoagulant RBC-MVs to the morbidity and mortality of extracellular histones in vivo requires further studies.
Monroe:Novo Nordisk A/S: Honoraria, Research Funding.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal